
Abstract
BACKGROUND AND PURPOSE: Whether Alzheimer disease (AD) represents exaggerated aging rather than a disease is controversial. Data about the effects of normal aging on the human brain are essential for clarifying this issue; however, whether coherent common patterns of regional morphologic brain changes emerge in the normal aged brain is unclear. Clarification of regional morphologic changes in the brain associated with normal aging and AD was sought using MR imaging.
METHODS: Ninety-two healthy volunteers and 26 mildly to moderately impaired patients with AD participated. Images were anatomically normalized, and voxel-by-voxel analyses were done.
RESULTS: In healthy volunteers, an age-related decline in the volume of the prefrontal cortex, insula, anterior cingulate gyrus, superior temporal gyrus, inferior parietal lobule, and precuneus was found. These decreases might contribute to the cognitive changes during normal aging. In patients with AD, a significant reduction of gray matter volume in the hippocampal formation and entorhinal cortex bilaterally was noted.
CONCLUSION: Morphologic changes associated with normal aging are clearly different from those seen with AD.
Alzheimer disease (AD) is the most common cause of dementia, demonstrating progressive decline in memory, language, and visuospatial abilities. Distinguishing AD from normal aging has been a recurring nosologic and diagnostic problem (1, 2). Whether AD represents exaggerated aging rather than a disease is a matter of controversy (3–12).
Not all researchers accept the notion that AD might be an exaggerated aging condition. Recent cognitive studies suggest that in truly normal aging, cognitive functions remain unimpaired over the life span, whereas sustained decline might represent a pathologic condition (1, 13, 14). In neuropathologic studies, Gomez-Isla et al (15) reported specific neuronal loss in the entorhinal cortex in persons with very mild AD and no change in the same region in the cognitively intact elderly. These observations imply that AD and normal aging are dichotomous.
Data about the effects of normal aging in the human brain are essential for clarifying this issue; however, whether coherent, common patterns of regional morphologic brain changes emerge in the normal aged brain is unclear. To date, regional differences in brain aging have not been investigated in a systematic manner in a large number of regions and specimens.
In our study, therefore, we first attempted to clarify regional morphologic changes in the brain associated with normal aging by using high-resolution MR images. We also attempted to clarify whether AD is an exaggerated aging process, in terms of comparison with patterns of regional morphologic brain changes in AD and normal aging. We applied a voxel-based morphometry technique for analyzing morphologic MR images to search the entire brain volume without subjectivity and an a priori hypothesis (16).
Methods
Research Subjects
Ninety-two healthy volunteers (90 right-handed, two left-handed) participated. The volunteers were divided into three age groups, each spanning two decades: a young group (31 men and 16 women; mean age, 28.5 years; SD, 6.0; range, 18–40 years), a middle-aged group (13 women and 10 men; mean age, 51.3 years; SD, 6.0; range, 41–60 years), and an aged group (12 women and 10 men; mean age, 71.2 years; SD, 5.8; range, 61–83 years). Volunteers having neurologic and psychiatric disorders, including alcoholism, substance abuse, atypical headache, head trauma with consciousness loss, asymptomatic cerebral infarction depicted by both T2- and T1-weighted MR imaging, hypertension, chronic lung disease, kidney disease, chronic hepatic disease, cancer, and diabetes mellitus, were excluded from this study. Neuropsychological tests, such as Mini-Mental State Examination, digit span, word learning and delayed recall, story recall, and Rey-Osterrieth complex figure test, were administered to the aged group to eliminate individuals with an early stage dementia (17). All healthy research subjects denied any use of psychominatic and other medications known to affect brain size, such as steroids.
Twenty-six right-handed patients (15 women and 11 men; mean age, 72.1 years; SD, 1.1; range, 59–79 years) meeting the criteria of the National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer's Disease and Related Disorders Associations for probable AD were recruited from our memory disturbance clinic. The mean age of the AD group was consistent with that of the healthy aged group. There was no evidence of concurrent cerebrovascular disease, except periventricular white matter intensities on T2-weighted or fluid-attenuated inversion recovery MR images in some patients. The patients with AD were mildly to moderately impaired, based on Mini-Mental State Examination scores (mean, 20.7; SD, 3.1; range, 16–26), and were considered free of depression on the basis of a score greater than 8 on the Hamilton Depression Rating Scale.
Written informed consent was obtained from all participants according to the declaration of Helsinki, as was the approval of our local ethics committee.
Brain MR Imaging
All MR studies were performed with a 1.0-T system. Three-dimensional volumetric acquisition of a T1-weighted gradient-echo sequence at 11.4/4.4/1 (TR/TE/excitation) produced a gapless series of contiguous, thin sagittal sections with the following parameters: flip angle, 15 degrees; acquisition matrix, 256 × 256; field of view, 31.5 cm; section thickness, 1.23 mm.
Image Analysis (Voxel-Based Morphometry)
The theory and algorithm of voxel-based morphometry have been well documented by Ashburner and Friston (16). Figure 1 presents a schema of voxel-based morphometry. The acquired MR images were reformatted to gapless, 2.3-mm-thick transaxial images. Image analysis was done using both statistical parametric mapping (SPM)99 and SPM96 (developed by KJ Friston et al, Wellcome Department of Cognitive Neurology, London, England) running on MATLAB (The Mathworks Inc, Sherborn, MA). Spatial normalization fitted each individual brain to a standard template brain (18) in 3D space, to correct for differences in brain size and shape and to facilitate intersubject averaging. Normalized MR images were then segmented into gray matter, white matter, cerebrospinal fluid, and other compartments by using a modified version of the clustering algorithm, the maximum likelihood “mixture model” algorithm (16, 19). The gray matter images were smoothed with a 12-mm, full-width half-maximum isotropic gaussian kernel to use the partial volume effect to create a spectrum of gray matter intensities. The gray matter intensities are equivalent to the weighted average of gray voxels located in the volume fixed by the smoothing kernel; therefore, regional intensities can be taken as equivalent to gray matter volumes (16).

Outline summary of voxel-based morphometry. Spatial normalization fitted each individual brain to a standard template brain in 3D space. Normalized MR images were then segmented into gray matter, white matter, cerebrospinal fluid, and other compartments. The gray matter images were smoothed with a 12-mm, full-width half-maximum isotropic gaussian kernel to use the partial volume effect to create a spectrum of gray matter intensities
Statistical Analysis
The processed images were analyzed using SPM96, which implements the “general linear model.” The global gray matter in the images, sex, and handedness were treated as confounding covariates. Proportional scaling was used to achieve global normalization of voxel values between images. In the SPM analysis for normal aging effects, we looked for all voxels where regional gray matter volume was significantly correlated with age by treating age as a covariate of interest. In the analysis for patients with AD, we studied differences in the gray matter between the young group versus the aged group and the young group versus the group with AD by using t statistics. The resulting sets of t values constituted statistical parametric maps: SPM (t). The SPM (t) were transformed to the unit normal distribution (SPM [Z]) and thresholded at P < .001. To correct for multiple nonindependent comparisons inherent in this analysis, the resulting foci were then characterized in terms of their spatial extent (k). This characterization is in terms of the probability that a region of the observed number or bigger voxels could have occurred by chance over the entire volume analyzed (Bonferroni correction, P < .05).
Results
Effects of Age on Regional Gray Matter Volume
Figure 2 shows the voxels with significant (P < .001) declines in gray matter volume with age. Six cortical areas were highlighted in both hemispheres: inferior frontal gyrus, insular cortex, cingulate gyrus, superior temporal gyrus, precuneus, and orbitofrontal cortex. Additionally, the left dorsolateral prefrontal cortex and the right inferior parietal lobule were highlighted in each hemisphere. The Table demonstrates these structures and their atlas coordinates (x, y, z) of the highest valued voxel in each area. There was no positive correlation with age and volume in the gray matter.

Negative correlations between age and regional gray matter volume in healthy volunteers. The SPM of the t statistics (after transformation to a SPM [Z] for this contrast) is displayed in a standard format as a maximum intensity projection viewed from the right-hand side and from the back and the top of the brain. The anatomic space corresponds to the atlas of Talairach and Tournoux (18). Significant reduction of regional gray matter volume is noted in the inferior frontal gyrus, the insular cortex, cingulate gyrus, superior temporal gyrus, precuneus, left dorsolateral prefrontal cortex, right inferior parietal lobule, and orbitofrontal cortex
Morphologic Changes in AD
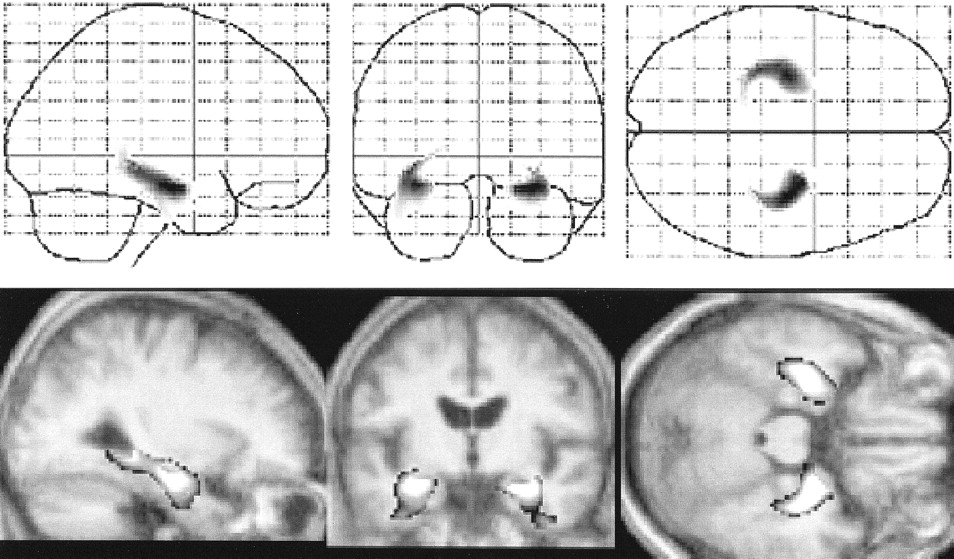
Figure 3 shows the voxels with significant (P < .001) decline in gray matter volume in patients with AD, in terms of an interaction between normal aging process and pathologic aging process. A significant reduction of gray matter volume in the bilateral hippocampal formation, entorhinal cortex, and parahippocampal cortex (z = 5.62, at +32,−10,−18 mm; z = 5.36, at −32,−12,−16 mm) was noted. There was no significant reduction of gray matter volumes in areas other than those demonstrated with normal aging effects.

Significant reduction of regional gray matter volumes in AD, which was noted in the bilateral hippocampal formation, entorhinal cortex, and parahippocampal cortex
Discussion
Normal Aging
Our results show a region-dependent vulnerability in gray matter with age in prefrontal cortex, insula, anterior cingulate gyrus, superior temporal gyrus, inferior parietal lobules, and the precuneus. All of these affected areas are polymodal and association cortices of the limbic system, believed to be related to cognitive processes that include attention, working memory, and the control of behavior. Damage to these areas, therefore, could lead to cognitive changes observed in the elderly. Several cognitive studies of healthy elderly individuals have suggested that the normal aging process should alter prefrontal cortex functions, including attention (20, 21). Rabbitt (21) was one of the first researchers to demonstrate experimentally that elderly subjects were less able to ignore task-irrelevant information. When aged people were tested with a battery of neuropsychological tests, frontal lobe deficits seemed to be a primary component of their cognitive impairments (22). The results of this study provide some support for these behavioral data.
Several postmortem and in vivo neuroimaging studies have suggested that prefrontal, entorhinal, and temporal cortices should be the most severely affected, whereas primary visual and somatosensory cortices might be more resistant to the influence of aging (12, 23–26). Postmortem studies have several limitations, such as technical and fixation artifacts, selection bias, and the influence of the illness and cause of death. In contrast, using in vivo brain imaging techniques can avoid these problems and provide an opportunity to examine normal brain morphology in healthy subjects. To date, the most comprehensive volumetric MR study that focused on regional changes in brain parenchyma was reported by Raz et al (25). Because of the voxel-based morphometric method applied in this study, it is difficult to directly compare our results with those from earlier volumetric MR studies using region-of-interest analysis. Although the latter approach has gained general acceptance, it has a limitation: the sample selection is dependent on the observer's a priori choice and hypothesis. It leaves large areas of the brain unexplored. Our voxel-based morphometric method is used to avoid subjectivity and dependence on a priori hypothesis and to adopt the principle of data-driven analysis. Raz et al also reported that the most substantial age-related decline was found in the volume of the prefrontal gray matter. There are, however, some discrepancies between their study and ours. Our study indicates an age-related decline in gray matter volume in the anterior cingulate gyri and insular cortices. Recent investigations with functional neuroimaging have revealed that the anterior cingulate gyrus should be involved in attentional processing, especially for divided attention (27, 28). Behavioral researchers have established that changes in attention functioning occur during aging (31, 32). A consistent finding from this research is that an age-related decline in performance is magnified when attention must be divided among several relevant stimuli (29–32). In this context, it is reasonable to speculate that the age-related change in the anterior cingulate gyri shown in this study should be related to these cognitive changes.
To our knowledge, neither an in vivo morphologic neuroimaging study nor a postmortem study has reported age-related changes in the insular cortex. The changes in the insular cortex have been ignored, because of limitations of the methodologies applied in earlier studies. Similar age-related changes in regional cerebral blood flow and regional glucose metabolism, including insular decline, were demonstrated by voxel-based analysis (33, 34). Several studies suggest that the insular cortex should be connected to a variety of paralimbic system and heteromodal association areas that are important in processing complex sensory information (35, 36). Therefore, atrophy of the insula cortex might contribute directly or indirectly to the cognitive deficits in the healthy elderly. Foundas et al (37) reported selective atrophy in the insula in patients with AD by using volumetric MR imaging. Some neuropathologic studies also suggested that the insular cortex should be involved in AD (38, 39). Our study, however, indicates that the atrophy in the insular cortex is a normal age-related change rather than a pathologic process of AD.
We could not find any age-related changes in the hippocampus. Animal and human postmortem studies reveal age-associated changes in the hippocampus; it is unlikely that in healthy humans this region is resistant to the influence of aging (24, 40, 41). One possible explanation should be considered. As mentioned earlier, we performed extensive neuropsychological tests to eliminate an early stage dementia. The inclusion of patients with early neurodegenerative disease in corrective studies of structural changes with age should significantly bias the result toward finding reduced hippocampal volume with age. Notably, West et al (42) reported no change in neuron number in several hippocampal subfields in normal aging. Sullivan et al (43) also reported that there was an age-related decline in MR imaging volumes of temporal lobe gray matter but not hippocampus. Our data support the hypothesis that hippocampal atrophy does not occur during strictly normal aging processes.
Increasing histopathologic evidence suggests that the alterations in white matter loss might represent the predominant neuroanatomic changes in normal human aging (44). Myelin loss might contribute to the cognitive deficits in aged individuals, because it is necessary for the rapid conduction of impulses along an axon. Therefore, changes in not only gray matter but also white matter would lead to the cognitive impairment in the elderly. Since we did not study changes in white matter volume associated with aging, further study is needed to clarify whether changes in white matter, rather than gray matter, contribute to the cognitive impairments in normal aging.
Morphologic Changes in AD
Our study indicates that morphologic changes associated with normal aging clearly differ from those associated with AD. Contrary to individuals with normal aging processes, patients with AD had decreased gray matter volumes in the medial temporal structure, hippocampal formation, entorhinal cortex, and parahippocampal gyrus, which are believed to constitute the long-term–memory system. Several cognitive studies have revealed that the impairment of episodic memory with relatively preserved semantic memory and working memory is a typical cognitive dysfunction in patients with mild AD (39, 45, 46). The morphologic changes in AD—altered medial temporal structures and relatively preserved prefrontal and medial frontal cortices—are concordant with the pattern of memory impairment observed in mild cases of AD. Many neuroimaging and neuropathologic studies have focused on changes of the medial temporal structure in patients with AD (5–12, 15, 24, 38, 39, 42). The neuropathologic studies have suggested that there is a selective vulnerability in AD (involving the CA1 zone of the hippocampus and layers 2 and 4 of the entorhinal cortex), whereas neuronal loss does not occur in these regions during the normal aging process (15, 42). Some MR studies of regional atrophy in AD suggest that the hippocampus is significantly decreased in volume, whereas other brain regions are less reliable markers of AD (37, 47). Our data indicate that the medial temporal structure should be the most vulnerable site in AD. That decreased gray matter volume in this region does not occur during normal aging is consistent with the hypothesis that AD and aging are not part of a continuous spectrum.
Conclusion
Using voxel-based morphometry, we found an age-related decline in the volume of the prefrontal cortex, insula, anterior cingulate gyrus, superior temporal gyrus, inferior parietal lobule, and precuneus in healthy volunteers. In patients with AD, a significant reduction of gray matter volume in the hippocampal formation and entorhinal cortex was noted. These results indicate that morphologic changes associated with normal aging differ from those that occur with AD. This supports a model in which AD and normal aging are not part of a continuum and can be differentiated.
Footnotes
1 Supported by the Research Grant for Nervous and Mental Disorders (10–1) from the Ministry of Health and Welfare.
2 Address reprint requests to Takashi Ohnishi, National Institute of Neuroscience, National Center of Neurology and Psychiatry, 4-1-1 Ogawa higashi, Kodaira City, Tokyo, Japan 187-0031.
References
- Received December 21, 2000.
- Accepted after revision April 3, 2001.
Location and peak of significant declines in gray matter volume with age
- Copyright © American Society of Neuroradiology
In this issue





{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- Structural Degeneration of the Nucleus basalis of Meynert in Mild Cognitive Impairment and Alzheimers Disease - Evidence from an MRI-based Meta-Analysis
- Relationship between finger movement characteristics and voxel-based specific regional analysis systems for Alzheimers disease
- Distinct changes in morphometric networks in aging versus Alzheimers disease dementia
- Network Curvature as a Hallmark of Brain Structural Connectivity
- Initial Experience in Using Continuous Arterial Spin-Labeled MR Imaging for Early Detection of Alzheimer Disease
- Age, Alzheimer disease, and brain structure
- A voxel based morphometry study on mild cognitive impairment
- Correction for Partial-Volume Effects on Brain Perfusion SPECT in Healthy Men
- Detection of grey matter loss in mild Alzheimer's disease with voxel based morphometry
- Longitudinal Evaluation of Both Morphologic and Functional Changes in the Same Individuals with Alzheimer's Disease