
Abstract
BACKGROUND AND PURPOSE: Prenatal drug exposure may influence the developing brain. Our aim was to study WM characteristics with DTI in children with prenatal opiate and polysubstance exposure and in controls. We assessed whether group differences in FA, DA, and DR could be found and related to cognitive function.
MATERIALS AND METHODS: The study was approved by a committee for medical research ethics. Parents signed an informed consent; children gave spoken consent. Our sample included 14 prenatally substance-exposed adopted children (5 girls; age range, 8.6–13.9 years; mean, 11.3 ± 1.7 years) and 14 control children (7 girls; age range, 9.0–10.2 years; mean, 9.8 ± 0.3 years). Tract-based spatial statistics were used to define a common WM skeleton for the sample, and FA was compared between groups throughout the skeleton, controlling for age and sex. Clusters of significant group differences ≥100 voxels (P <. 05) were identified. FA, DA, and DR within clusters were correlated with cognitive function.
RESULTS: Ten clusters of FA group differences, mostly in central, posterior, and inferior parts of the brain, were identified (P <. 05), showing lower FA in substance-exposed children. FA and DA correlated positively and DR, negatively with cognitive function across groups.
CONCLUSIONS: Prenatally substance-exposed children exhibited lower FA in restricted areas of WM, mostly relatively central, inferior, and posterior, where myelination occurs early in development. Myelin in these areas may be particularly vulnerable to prenatal substance exposure. FA and DR related moderately to cognitive function. Potential confounding factors existed and were considered.
Abbreviations
- ADHD
- attention deficit/hyperactivity disorder
- ANOVA
- analysis of variance
- AS
- Asperger syndrome
- ATRL
- anterior thalamic radiation, left
- ATRR
- anterior thalamic radiation, right
- CC
- corpus callosum
- CGL
- cingulum gyrus, left
- CGR
- cingulum gyrus, right
- CHL
- cingulum hippocampal part, left
- CHR
- cingulum hippocampal part, right
- CNS
- central nervous system
- CSTL
- corticospinal tract, left
- CSTR
- corticospinal tract, right
- DA
- axial diffusion
- DR
- radial diffusion
- DTI
- diffusion tensor imaging
- FA
- fractional anisotropy
- FASD
- fetal alcohol spectrum disorders
- FFD
- freedom from distractibility
- Fmaj
- forceps major
- Fmin
- forceps minor
- FMRIB
- Functional MRI of the Brain
- GCI
- general cognitive index
- GLM
- general linear model
- ILF
- inferior longitudinal fasciculus
- ILFL
- inferior longitudinal fasciculus, left
- ILFR
- inferior longitudinal fasciculus, right
- IQ
- intelligence quotient
- L
- left
- PPS
- Perceptual Performance Scale
- R
- right
- SES
- socioeconomic status
- SLF
- superior longitudinal fasciculus
- SLFL
- superior longitudinal fasciculus, left
- SLFR
- superior longitudinal fasciculus, right
- UF
- uncinate fasciculus
- UFL
- uncinate fasciculus, left
- UFR
- uncinate fasciculus, right
- WISC-R
- Wechsler Intelligence Scale for Children-Revised
- WM
- white matter
Drugs of abuse can potentially affect mitosis, migration, and survival of cells in the developing CNS. Children born to substance-abusing women have increased risk of regulatory and neuropsychological difficulties,1–4 and neuroanatomic volumetric differences between exposed children and controls have been documented for alcohol,5 methamphetamine, and heroin.6–8 In the formation of the human CNS, neurons arise first, followed by astrocytes, and then oligodendrocytes.9 The latter form myelin, the insulating layer around axons, which enables speedy and synchronized transmission of nerve signals. Brain myelination begins relatively late, mostly after midgestation, develops rapidly in infancy, and continues at a slower pace well into adulthood.9 However, much of the foundation of myelination is cast in utero, so one should examine potentially relevant factors at this stage.
Little is known about the effects of different drugs on developing WM, but WM abnormalities, particularly in the posterior CC, have been well documented by structural MR imaging in FASD.5,10 It has also been reported that children born to mothers with opiate and polysubstance abuse have smaller neuroanatomic volumes, including WM volume.8 This finding may be linked to studies showing opiate-induced apoptosis in neurons and microglia.11 Less is known about mechanisms by which opiates and other drugs regulate oligodendrocytes, but these cells also express specific opiate receptors and can respond to changes in the level of stimulation.12 Opioid receptor activation can regulate cellular development, so one could speculate that chronic opiate exposure might cause lasting changes in neural function by disrupting gliogenesis.13
DTI provides a measure of microstructural WM characteristics by using the restricted water diffusion in biologic brain tissues. FA is an intravoxel index of the degree of diffusion anisotropy. Several features contribute to FA, including degree of myelination and axonal integrity.14,15 FA consists of components measured DA and DR to the principal diffusion direction. Three DTI studies of WM characteristics in FASD have been conducted, and all reported effects in the posterior CC.10,16,17 No DTI studies have been reported on effects of prenatal exposure to drugs of abuse other than alcohol.
The present study targets FA, DA, and DR of WM in children with prenatal polysubstance, predominantly opiate, exposure. To our knowledge, there are no previous studies in this area, so the present study is explorative and hypotheses are tentative. To the extent that such exposure disrupts myelination, we expected lower FA and higher DR in exposed children. Myelination begins prenatally9 and appears to progress in an orderly manner during infancy from inferior to superior, posterior to anterior, and central to peripheral regions.18 We, therefore, hypothesized that the largest effects of prenatal substance exposure would be found on relatively central, inferior, and posterior WM. Finally, on the basis of the important role of cerebral WM in cognitive function,15 we expected FA and DA to correlate positively and DR to correlate negatively with IQ and FFD as measured by the WISC-R.19
Materials and Methods
Sample
The present sample is drawn from a longitudinal project on the development of children born to mothers who abused drugs during pregnancy, further details of which are described elsewhere.3,4,8 The biologic mothers of the substance-exposed children were referred to the perinatal risk project because of concerns about substance abuse by medical or social staff at the municipal health service. Control children without known risk of biologic mothers with a history of alcohol or illicit drug abuse were recruited from local maternal and child health centers. Fourteen children prenatally exposed to drugs of abuse without a diagnosis of FASD and 14 controls were included in the present study. Sample characteristics are presented in Table 1.
Sample characteristics of the 2 groups
One-way ANOVA showed significant group differences in several characteristics. Because the inclusion of 2 prematurely born children (see below) may have disproportionately increased group differences, analyses were also repeated without these children. All significant group differences remained. After exclusion, in the substance-exposed group, mean gestational age was 39.0 ± 1.3 weeks; birth weight, 3158 ± 595 g; birth length, 48.3 cm (SD, 2.6 cm); and head circumference, 34.5 ± 1.6 cm. The biologic mothers of 10 of the substance-exposed children reported opiates as their main drug of choice. Nine of these women reported use of a range of 1–5 other substances, including benzodiazepines, neuroleptics, cannabis, alcohol, cocaine, and amphetamine. Of the 4 mothers not reporting heroin use, 1 reported cocaine; 2, psychopharmacologic substances; and 1, alcohol as the main substance of choice, in addition to various other substances, including benzodiazepines, neuroleptics, amphetamine, and alcohol. Twelve of the substance-exposed children had neonatal abstinence syndrome20 symptoms, and 10 were medicated during the first 8 weeks after birth. All biologic mothers of the exposed group reported smoking tobacco during pregnancy. One control mother reported infrequent cigarette smoking. All substance-exposed children moved from their biologic mothers within 6 weeks after birth, were placed in permanent foster care within their first year of life, and were later adopted by the same foster parents.
Among the opiate-exposed children, 1 was born with a myelomeningocele. All brain scans were inspected by a neuroradiologist. They were examined for anatomic abnormalities, including congenital changes, and the neuroradiologist looked for signal-intensity differences/pathology in white and gray matter and examined CSF spaces. No signs of cerebral pathology were found. This was also the case with the scans of the child with myelomeningocele. There was no sign of intracerebral pathology, either hydrocephalus or any Chiari malformation. Another of the opiate-exposed children had been diagnosed with AS. We decided to include both children. Maternal opiate addiction may be associated with significant prenatal risk both because of direct teratogenic factors and poor health and dietary conditions. Myelomeningocele may be associated with inadequate maternal folic acid intake, low maternal vitamin B12 status, and exposure to some teratogens; and approximately 25% of the incidence of AS is associated with pre- or perinatal risk.21,22 Prenatal substance exposure is also associated with heightened risk of premature birth23 and ADHD.4 Two of the opiate-exposed children were born prematurely (gestational age < 37 weeks, namely 31 and 34 weeks), and 4 of the substance-exposed children in the present sample had an ADHD diagnosis, for which they were medicated. The study (including both substance-exposed and control children) was approved by the regional committee for medical research ethics for South Norway. All parents signed informed consent. The children were given adapted information (information in a simplified language), and all gave spoken consent.
As part of previous follow-ups, the sample has undergone behavioral assessments.2–4 For all except 1 child, the WISC-R19 was administered at a mean age of 8.63 ± 0.71 years. The WISC-R is a general ability measure comprising 12 subtests. Total IQ is calculated from all subtests, while the FFD factor is comprised of the Arithmetic, Digit Span, and Coding subtests. It has been suggested that this factor reflects resistance to the effects of distracting stimuli, and executive and short term memory processes involved in planning, monitoring and evaluating task performance.24
DTI Acquisition and Processing
A Sonata 1.5T MR imaging scanner (Siemens, Erlangen, Germany) with an 8-channel head coil was used. A single-shot echo-planar imaging−based 6-direction diffusion-weighted sequence was used with 40 axial sections (section thickness, 2.6 mm; FOV, 230 mm; in-plane pixel size, 1.7969 × 1.7969 mm; TR/TE/ flip angle, 6100 ms/85 ms/90°). Two-to-4 successive runs were used, with 1 non-diffusion-weighted (b = 0) image in addition to 6 diffusion-weighted images obtained per acquisition. The volumes were eddy current− and motion-corrected relative to the first b = 0 volume by using the Diffusion Toolbox of FMRIB, part of FMRIB Software Library (http://www.fmrib.ox.ac.uk/fsl/).25 To preserve the orientational information after motion correction, we reoriented the B matrix accordingly by applying the resulting rotations from the motion-correction procedure.26 The acquisitions were averaged before extracting nonbrain tissue (skull, CSF, etc) by using the Brain Extraction Tool.27 The Diffusion Toolbox of FMRIB was used to fit a diffusion tensor model to the data at each voxel. Voxelwise values of FA as well as DA (λ1) and DR ([λ2 + λ3] / 2) to the principal diffusion direction were calculated.
Statistical Analyses
Voxelwise statistical analysis of the FA data were carried out by using Tract-Based Spatial Statistics,28 part of the FMRIB Software Library.24 All subjects' FA data were aligned into a common space by using nonlinear registration.29–31 Every participant was registered to every other participant, and the most typical was chosen as the target (ie, the target image that minimized the amount of warping required for all other subjects to align to it).28 The mean FA image was created and thinned to create a mean FA skeleton representing the centers of all tracts common to the group. The skeleton was thresholded at FA > 0.25 and included 131,198 1-mm3 voxels. For a discussion of the choice of thresholding procedure, see the Appendix.
The tract invariant skeleton for each participant was used to model group effects in a GLM, with age and sex as covariates. Cluster-size thresholding was carried out on the resulting statistical maps by clustering contiguous voxels by means of 26-neighbor connectivity in the skeleton with a cluster-forming threshold of P < .05 for the 2 contrasts (Exposed > Controls, Controls > Exposed). Due to the small number of this rare sample, stringent permutation-based corrections were deemed too strict. Instead, to ensure stability of effects, we selected only clusters with ≥100 voxels for further analyses, and reliability within these were assessed by split-half analysis. The FA, DR, and DA values within identified clusters were subjected to the split-half analysis, where each sample was divided into halves matched with respect to age and sex, and analysis was run on the 2 subsamples. Furthermore, group differences in FA, DR, and DA values within identified clusters were also subjected to analysis with the substance-exposed sample limited to only children exposed to heroin and to only children exposed to heroin born at term and not having a diagnosis of myelomeningocele or AS. To explore the regional distribution of the effects from the whole brain analyses not restricted by the cluster size threshold of ≥100 voxels, we counted all voxels yielding a statistically significant (P < .05) group difference in FA within probabilistically defined fiber tracts.32–34 This method has previously been used in conjunction with and in addition to cluster-size inference.35
Results
Group Differences
A cluster-forming threshold of P < .05 identified 10 clusters of ≥100 voxels for the Controls > Exposed contrast and none for the Exposed > Controls contrast. The clusters are shown in Fig 1 and On-line Fig 1, and their size and Montreal Neurological Institute coordinates are listed in On-line Table 1. In general, clusters were found in relatively central, posterior, and inferior WM (see Table 2 for a short anatomic description). One exception was a right-sided cluster in an anterior part of the Fmin. Results for group differences in DA and DR, respectively, from a GLM with age and sex as covariates, are shown in Table 2, and group mean values are shown in Fig 2 . Differences in DR, more than DA, contributed to the lower FA in the substance-exposed children. In all cases of significant differences, DA was lower and DR was higher in exposed children. The group differences within these clusters were upheld when the sample groups were split in half (matched with respect to age and sex) and the analysis was rerun on the 2 subsamples. The results of the split half analysis are shown in On-line Table 2. For 7 of 10 clusters, the group effects were upheld at P < .05 in both halves of the sample, despite the substantial reduction in sample size. The 3 other clusters, I, H, and J, showed group effects at P < .05 in subsample A and around P ≤ .10 in subsample B in the split-half analysis. The group differences in FA, DR, and DA values within identified clusters were also subjected to reanalysis with the substance-exposed sample limited to only children either exposed to heroin or exposed to heroin, born at term, and not having a diagnosis of myelomeningocele or AS. In both cases, all group differences in FA and DR in the clusters remained significant (P < .05). The results of these analyses are shown in On-line Table 3.

Clusters of voxels (≥ 100) with significant (P < .05) group differences in FA are shown in red-yellow as they appear in the WM skeleton shown in green. For all clusters, FA was lower in the prenatally substance-exposed children.
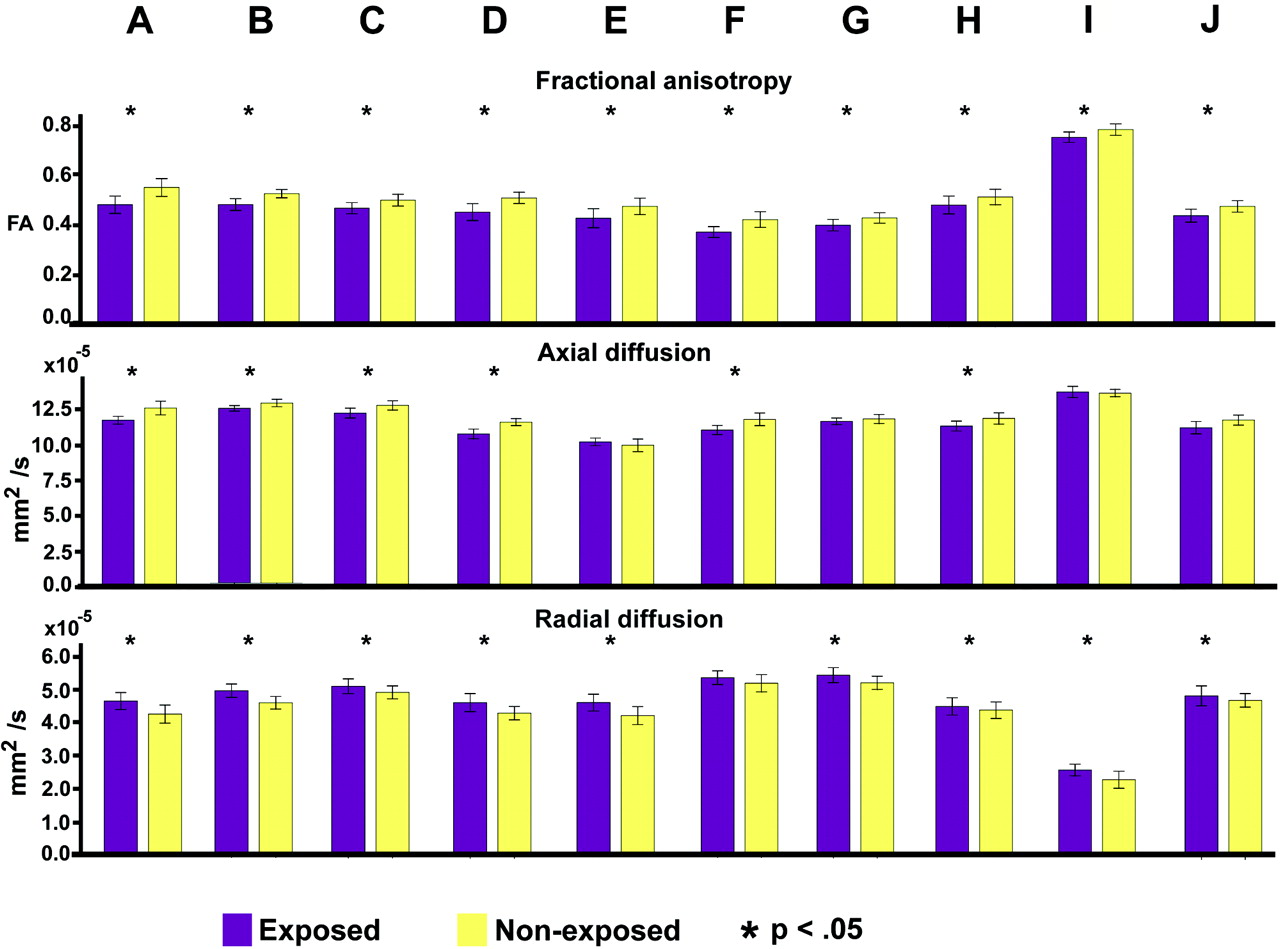
Group mean values and standard errors of the mean in the 10 identified clusters for FA, DA, and DR in substance-exposed and control children.
Results for group differences in FA, DA, and DR from a GLM analysis, with age and gender entered as covariates
Tract-wise percentages of voxels showing significant (P < .05) group differences in FA are shown in On-line Fig 2. Only the ILFR showed group differences in mean FA (F = 4.327, P = .048). A trend was also observed for the SLFL (F = 3.666, P = .068). However, it is evident that most tracts (8 of 9) showed a larger proportion of significant voxels in the Nonexposed > Exposed contrast than in the Exposed > Nonexposed contrast, as expected from the cluster size inference. These effects were distributed unevenly across tracts, suggesting some degree of anatomic specificity. The most affected tract was the ILF, followed by the SLF.
Correlations between Cognitive Variables and FA and Its Constituents
Five clusters correlated with cognitive function when the effects of age and sex were accounted for statistically: In the right deep temporal WM (cluster C), FA correlated positively with FFD (r = 0.42, P = .039) and DR correlated negatively with IQ (r = −0.46, P = .021) and FFD (r = −0.56, P = .004). The latter correlation was also seen with marginal significance for the opposite hemisphere counterpart (cluster B) of this cluster: r = 0.40, P = .051. DA in the superior division of the left lateral occipital area (cluster D) correlated positively with FFD (r = 0.44, P = .026). DR in the left inferior temporal gyrus (E) was negatively related to FFD (r = −0.40, P = .045), and FA in the right occipital/fusiform temporal gyrus (F) was positively related to IQ (r = 0. 44, P = .028). Finally, both FA and DR in the right prefrontal part of the Fmin (cluster J) correlated with IQ (r = 0.48, P = .016 and r = −0.49, P = .013 respectively). In a follow-up analysis, the observed significant correlations were rerun when the sample was split by group (Exposed versus Controls) to see whether these were primarily found in one group. This was not the case—with the exception of a negative correlation (r = −0.58, P = .048) between DR and IQ in the Fmin (cluster J) in the control group, the above correlations were nonsignificant (P < .05) within groups.
Discussion
This study showed differences in FA between children prenatally exposed to drugs of abuse, primarily opiates, and control children in regions of the WM of the brain. All identified clusters of differences showed lower FA in substance-exposed children. With the exception of a cluster in the Fmin, all effects were located in relatively central, posterior, and inferior parts of the brain. This is where myelination occurs relatively early in CNS development,18,36 and differences in these areas can be interpreted as corresponding to agents of early influence, though a number of potential confounds must also be considered. There were, to a somewhat greater extent, more group differences in DR than in DA across clusters. The exact neurobiologic principles governing radial and axial diffusivity are unknown, but DR has been related to the integrity and thickness of myelin sheets covering the axons,37,38 whereas DA may index axonal integrity.39
A recent autopsy study of adult opiate abusers did not find evidence for altered apoptotic pathways,40 but in keeping with histologic studies,11–13 opiate exposure may still alter fetal neuronal and glial development. One may thus speculate that substance exposure has a negative effect on myelin development, here reflected by increased radial diffusivity, though the exact molecular basis is unknown. Prenatal alcohol exposure may yield decreased expression of myelin basic protein41 and alter protein trafficking in astrocytes.42 Unfortunately, we were unable to find further studies that more specifically link ultrastructural changes in prenatal opiate and polysubstance exposure to the present DTI findings. The consequences of opiate use in pregnancy have not been well characterized, and animal models on fetal opiate exposure incorporating histologic, imaging, and behavioral observations would be most useful. Only a relatively low proportion of WM becomes myelinated in utero. No effects were observed in the early-myelinating brain stem, suggesting that there is not a straightforward correspondence between the timing of myelination, prenatal substance exposure, and later DTI effects. Additional factors governing localization of effect sites are unknown, but the regional distribution of different types of opiate receptors, for example, is known to vary,43 and this variation may be a possible source of influence. Some of the observed effects lie within areas known to myelinate prenatally (eg, the posterior limb of the internal capsule),44 whereas others are in areas where myelin is not yet seen at birth. However, both biochemical detection of myelin-associated lipids and proteins and increases in FA are known to precede the histologic detection of myelin.45,46 Hence, it is possible that mechanisms functioning before myelination could be affected by drug exposure in utero and ultimately affect the development of myelin.
Any neurobiologic inferences based on DTI-derived measures must be made with caution. FA is also influenced by crossing fibers and axon diameters, and increases in both can lead to increased DR, while potentially still being beneficial to cognitive function.47 The functional significance of the present findings can thus not be determined with certainty. Some of the presently identified clusters of group difference correlated with IQ and/or FFD in a predictable way in the total sample (ie, higher FA and DA correlated positively with cognitive performance and DR correlated negatively). The observed significant correlations in the total sample were, with the exception of a negative correlation between DR in a cluster in the Fmin and IQ in the control group, nonsignificant when the sample was split into controls and exposed children, suggesting that between-group differences, rather than individual differences within the group, drive these relationships. Sowell et al,10 on the other hand, reported that FA of the CC splenium correlated with visuomotor integration only in an FASD group and not in controls. Both measures and the nature of the exposure differed across these studies, so they cannot be directly compared. It is interesting, however, that the present group effects in deep temporal WM appear to correspond to 1 of the main sites of group difference identified in the study by Sowell et al. Sowell et al also found that gross structural WM volume was lower in some but not all regions in which FA was lower. Regional WM volume was not measured in the present study, but a previous report from the same sample showed generally smaller neuroanatomic volumes in the exposed children, including gross WM volume.8
The ILF and SLF showed the greatest proportion of voxels with lower FA in substance-exposed children. These are 2 of the most prominent association tracts bridging frontal, temporal, and parietal regions. In young healthy persons, FA in these tracts has previously been shown by Turken et al48 to correlate positively with scores on the Digit Symbol Substitution Test, which is also a part of WISC-R and the FFD score. The presently identified bilateral temporal effects in the ILF correspond to areas identified by Turken et al. They point out that these WM tracts support frontoposterior network interactions and that they are presumably critical for relaying the analysis of visual features to centers involved in control of visual scanning and response selection. The interaction of posterior association areas involved in perceptual representations and transformation and frontal areas involved in attentional control and regulation of actions may thus be compromised by prenatal drug exposure, with reduced myelination of these tracts.
The present study has a number of limitations. First and foremost, the exposure group studied is rare and, therefore, small. This feature warrants caution in judging the stability of the effects. However, the split-half analysis did support stability of the observed effects. All significant group differences were in the same (and predicted) direction. While specific mechanisms of action have been identified for different drugs, including opiates, there is also neuroimaging evidence to suggest that there may be some common mechanisms affected by prenatal exposure to different drugs.49 The specificity of effects of different drugs cannot be well assessed in this small study. However, for the entire observed group, differences in FA and DR remained significant when the substance-exposed sample was limited to the children exposed to heroin. Furthermore, all the observed group differences in FA and DR remained significant when the substance-exposed group was further reduced to the few children (n = 6) exposed to heroin and not born prematurely or having diagnosed AS or myelomeningocele.
Another limitation is that we cannot control for possible genetic differences between the groups studied, because differences in FA and FA-cognitive performance correlations may also be influenced by this.50,51 Other possible confounds include poor maternal nutrition and health. Furthermore and potentially related to all of the above, there is an increased incidence of other difficulties, including ADHD symptoms, in the substance-exposed group.3,8 All of these factors may affect both brain and cognition. Hence, we cannot attribute group differences among substance-exposed and control children and their correlation with cognitive function to the direct influence of substances on CNS development alone. This does not, however, make the observed differences and associations less important to further research.
Conclusions
The present study showed that in children prenatally exposed to drugs, primarily opiates, FA was lower and DR higher, relative to controls. These differences were mostly seen in relatively deep, central, and posterior parts of WM tracts and were moderately related to cognitive performance across groups. These are areas where myelination starts relatively early and, to some extent, prenatally. In correspondence with findings from histologic studies, opiate exposure in utero may have neurotoxic as well as gliotoxic effects on WM development. However, one cannot conclude with certainty, given potential confounding factors such as genetic differences and maternal nutrition and health. There is, therefore, a need to research effects of prenatal substance exposure in additional samples.
Footnotes
-
This work was support by the Norwegian Research Council and the Institute of Psychology at the University of Oslo.
Indicates article with supplemental on-line tables.
Indicates article with supplemental on-line figures.www.ajnr.org
Indicates open access to non-subscribers at www.ajnr.org
References
- Received September 23, 2009.
- Accepted after revision October 13, 2009.
- Copyright © American Society of Neuroradiology
In this issue





{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- A Prospective Multi-Institutional Study Comparing the Brain Development in the Third Trimester between Opioid-Exposed and Nonexposed Fetuses Using Advanced Fetal MR Imaging Techniques
- DTI of Opioid-Exposed Fetuses Using ComBat Harmonization: A Bi-Institutional Study
- Consequences of Parental Opioid Exposure on Neurophysiology, Behavior, and Health in the Next Generations
- White Matter Injury and Structural Anomalies in Infants with Prenatal Opioid Exposure
- Multimodal imaging of the self-regulating developing brain